

PAPHIOPEDILUM PHILIPPINENSE has been in cultivation for over 100 years. In the early years, it was rediscovered, renamed and redescribed multiple times, leading to a conclusion by some that this is a highly variable species in color, shape and size. In the literature, it is often not described separately, but lumped in as a member of a larger orchid subgroup, which often includes rothschildianum, sanderianum, stonei, praestans, haynaldianum and lowii, as if the authors did not wish to expend the effort on an uninteresting or confusing species. Yet, the longevity of philippinense in cultivation belies its lack of documentation in the literature. It is a foundation plant; it is the backbone of many successful Paphiopedilum breeding programs for vigor, floriferousness and marketability. In addition, different aspects of philippinense have influenced traits from breeding within the species and in some of our most popular hybrids.

The Philippine Paphiopedilum was described as Cypripedium philippinense by Heinrich Gustav Reichenbach in Bonplandia in 1862 (Reichenbach 1862) before reclassification as Paphiopedilum philippinense by Stein in Stein’s Orchideenbuch in 1892 (Stein 1892). It has also been named Cypripedium laevigatum (Hooker 1865), Selenipedium laevigatum (May 1885), Cypripedium cannartianum hort. (Linden 1887), Paphiopedilum laevigatum (Pfitzer 1888) and Cordula philippinensis (Rolfe 1912). It is currently known as philippinense.

INFRAGENERIC CLASSIFICATION
The genus Paphiopedilum was originally categorized taxonomically through morphological characteristics, resulting in philippinense being placed in multiple differently named subgenera. For example, Pfizer placed it in Polyantha in 1894, before moving it to Anotopedilum in 1903. Brieger renamed it back to Polyantha in 1971 before Cribb moved it to Paphiopedilum Section Coryopedilum in the first edition of the Genus Paphiopedilum (Cribb 1987). This continued movement of the genus Paphiopedilum has led to some confusion in hybrid names and genealogy over the years, when they were based purely on morphological characteristics.
Cox et al. (1997) were the first to show common ancestry or monophyly among the slipper orchids based on molecular sequencing of nuclear DNA instead of morphology. They also showed that, genetically, paphiopedilums were more closely related to each other than to Selenipedium, Cypripedium, Phragmipedium or Mexipedium. This came as no surprise to taxonomists. However, the relationship within the sections or subgenera of paphiopedilums, depending on classification system, is much more dubious. In the second edition of the Genus Paphiopedilum, Cribb revised his Paphiopedilum classification to include genomic sequencing data from the earlier study (Cribb 1998).
The current classification by Cribb diviides the genus Paphiopedilum into three subgenera: Parvisepalum, Brachypetalum and Paphiopedilum. He further divided subgenus Paphiopedilum into sections Barbata, Cocholopetalum, Pardalopetalum and Coryopedilum. Cribb places Paphiopedilum philippinense in subgenus Paphiopedilum Section Coryopedilum. Unfortunately, the internet has made continued usage of older nomenclature possible, and Paphiopedilum philippinense is most often found in subgenus Polyantha as attributed to Karasawa and Saito (1982) in their classification.
Chochai et al. (2012) examined monophyly within paphiopedilums incorporating nuclear and plastid DNA and the categorization of Cribb (1998). In the study, it could be claimed with confidence that in Paphiopedilum, the subgenera Parvisepalum, Brachypetalum and Paphiopedilum formed three independent, monophyletic clades or groups with a common ancestor. Within subgenus Paphiopedilum, Section Coryopedilum is most closely related to Section Pardalopetalum, and most distantly related to Section Paphiopedilum (Chochai et al. 2012). To date, there have been no genetic studies examining the genetic distance between different varieties of philippinense. Thus, the varietal designations are based on morphology alone.
[1] Paphiopedilum philippinense ‘Carola’ AM/AOS best fits the concept of var. philippinense. Exhibitors: Armando and Carolina Betancourt.

[2] Awarded as phippinense ‘Hector’ FCC/ AOS, this cultivar exemplifies the concept of var. roebelenii with its long, twisted, darkly colored petals that drop nearly straight down. Exhibited by Ramon de los Santos

[3] Paphiopedilum philippinense var. compactum ‘Windy Hill’s Sprite’ CHM/AOS. Exhibitor: Marilyn LeDoux.

[4] Awarded as Paphiopedilum laevigatum ‘Mikado’ HCC/AOS in 1981, this is now considered synonymous with philippinense var. philippinense. Exhibitor: Walter Bertsch.

DISTRIBUTION
Paphiopedilum philippinense is found throughout the Philippines and extending southwest to the northeastern corner of the island of Borneo. This includes the islands off the coast of northern Borneo. It is found from sea level up to 1,640 feet (500 m) on limestone outcroppings, in leaf mulch, and on trees, often in completely exposed spaces. In contrast, the range of philippinense var. roebelenii is much smaller, endemic to the island of Luzon in the Philippines. Paphiopedilum philippinense plants require bright light, frequent watering, well-drained media and a diurnal difference of over 11 F (6 C) to induce flowering. In the Philippines, blooming occurs summer through fall, and in the United States, the onset of cooler temperatures in the fall induces flowering from January to April.
FORMS
Paphiopedilum philippinense var. philippinense is a medium-tolarge, warm-growing Paphiopedilum. It has large solid-green leaves approximately 2 inches (5 cm) wide that need to reach 7.1 inches (18 cm) in length prior to blooming, but which can reach up to around 20 inches (50 cm) in length. It is characterized by simultaneous blooming of multiple flowers on an inflorescence with long thin, curling petals.
[5] Paphiopedilum philippinense f. alboflavum ‘Sunset Valley Orchids’ AM/AOS. Exhibitor: Fred Clarke.

[6] Awarded as philippinense var. album ‘Albino Beauty’ AM/AOS, this best fits var. alboviride. Exhibitor: Orchid Inn, Ltd.

The flowers of typical examples of philippinense var. philippinense have white sepals striped maroon, thin, curling, maroon petals, and a yellow pouch. In most cases, the petals are long, thin and gently twisted, and the dorsal sepal is broader than the synsepal. The petal length can be highly variable. Though it was known for many years as Paphiopedilum laevigatum as described by Bateman in 1865, the descriptions matched those of the modern-day Paphiopedilum philippinense var. philippinense, leading to the official designation of laevigatum as a synonym for philippinense var. philippinense. However, in Asia, the Taiwan Paphiopedilum Society recognizes a variety of philippinense with short petals and only a slight twist as var. laevigatum. The sepals of Paphiopedilum philippinense var. laevigatum are white with red stripes, the petals are yellow with red, and the pouch is yellow. The petals are straighter, smoother, and extend almost horizontally to the side. Plants are also smaller than var. philippinense and var. roebelenii, and, in some cases, diminutive enough to be considered miniature but generally larger than var. compactum (also now a synonym of var. philippinense) described below.
Röbelen’s Paphiopedilum was described as Cypripedium roebelenii by Reichenbach in the Gardeners’ Chronicle in 1883, where he called it a rival species to philippinense (Reichenbach 1883). It has also been described as Cypripedium philippinense var. roebelenii (Kent 1889) and Paphiopedilum roebelenii by Pfitzer in 1894. It was moved to a varietal form of philippinense in the Genus Paphiopedilum by Cribb (1987), who did not consider it substantially different enough to merit species rank; it is currently known as Paphiopedilum philippinense var. roebelenii.
Paphiopedilum philippinense var. roebelenii was originally described in the Gardeners’ Chronicle as differing from Paphiopedilum philippinense as “[t]he leaves are narrower. The peduncle is more hairy…The hooded staminode has a small apiculus between its two anterior teeth” (Reichenbach 1883). It was also determined by horticulturalists and breeders at some point in the last 100 years to be darker in color than var. philippinense. A larger plant, the petals are generally longer and the majority of plants, over 90%, have petals that hang straight down, whereas the petals of var. philippinense extend to the side before curving downwards. Because of the difference in angle of the petals between var. philippinense and var. roebelenii, when comparing the horizontal natural spread of flowers, var. philippinense is invariably wider than var. roebelenii and the corollary is also true; var. roebelenii exhibits greater vertical natural spread than var. philippinense. The roebelenii variety is also known to have a differently colored and differently shaped staminodal shield from var. philippinense. The original description in the Gardeners’ Chronicle in 1883 states the “staminode has a small apiculus between its two anterior teeth,” whereas the original description of the staminode for var. philippinense in Bonplandia in 1862 by Reichenbach describes it as “staminodio cordiformi,” which, when translated from Latin, becomes “staminode cordiform” suggesting a heart-shaped staminode with no mention of apiculi or anterior teeth (Reichenbach 1862). Fowlie has a low resolution photograph on orchidspecies. com of a purported Paphiopedilum roebelenii staminode, showing a small protrusion or apiculus between the two anterior teeth but qualifies the photograph with the statement that the staminode only differs slightly from that of var. philippinense. Thus, for hobbyists and the majority of breeders, the major distinguishing morphological characters distinguishing var. philippinense and var. roebelenii is the angle of the petal extension and the length of the petals.
An albinistic form was first described by Valmayor & Tiu in 1984 as f. album, although the name was published invalidly and therefore has no taxonomic standing. In 2000, Olaf Gruss described the yellow and white forms as f. alboflavum, and in 2016 described the yellow and green forms as f. aureoviride (Gruss 2016). The background color of both forms is white and they differ in the color of the spots, stripes and pouch markings; yellow in f. alboflavum and green in f. aureoviride. Both Dave Sorokowsky of Paph Paradise and Sam Tsui of Orchid Inn, who use Paphiopedilum philippinense in their breeding programs, consider all three named forms to be synonyms of the albino form.
[7] Paphiopedilum philippinense ‘Penn Valley’ AM/AOS; exhibitor: W.W. Wilson.

[8] Paphiopedilum philippinense ‘Seven Samurai’ AM/AOS; exhibitor: Sam Tsui, Orchid Inn, Ltd.

[9] Paphiopedilum philippinense ‘Sam’s Choice’ FCC/AOS; exhibitor: Sam Tsui, Orchid Inn, Ltd.

A compact variety, Paphiopedilum philippinense var. compactum, was first described in Die Orchidee (Gruss et al. 2008). It was originally named Paphiopedilum palawanense when it was discovered some years earlier on Palawan, a Philippine island between Luzon and Borneo. However, the authors subsequently realized that the epithet palawanense had already been used in 1901 in “Sander’s Orchid Guide” for the hybrid of Paphiopedilum rothschildianum and Paphiopedilum dayanum, which is currently registered as Paphiopedilum Kimballianum. Thus, Gruss et al. chose to designate these small, compact specimens as a variety of philippinense. For a few years prior to the official description in Die Orchidee, var. compactum was known as Paphiopedilum philippinense var. palawanense and frequently conflated with var. laevigatum. Today, the prior color forms and var. compactum are all parts of var. philippinense.
Paphiopedilum philippinense var. compactum plants are much smaller than the conventional variety, with leaves of similar width but half the length, giving an overall plant width of about 12 inches (30 cm) or less, making it half the size of var. philippinense. Plants also tend to have fewer flowers per inflorescence, averaging 2–4, compared to the 4–6 flowers expected of typical var. philippinense. The plant also blooms earlier from flask. Compact forms occur in both conventional coloring and albino plants. So-called Paphiopedilum philippinense var. compactum f. aureoviride (small, compact yellow-andgreen forms) combine the tendency toward smaller size in albino plants with the compact variety to give the smallest philippinense yet.
SPECIES AWARDS
There have been 143 awards given to philippinense, with plants still being awarded every year. It is also the primary parent of 214 registered hybrids. Of these 143, 34 awards (24%) are specifically to var. roebelenii; spread evenly among quality and cultural awards. As of this writing in early 2023, there have been three AOS awards to var. compactum. Indeed, two awards were given in 2022 alone. This shows that despite being a long-cultivated species, philippinense continues to maintain its popularity and to develop in new directions.
Although philippinense var. roebelenii is distinct from var. philippinense based on its range, size, petal length, petal orientation and coloration, intermingling of var. roebelenii and var. philippinense for over 100 years leads to a distinct challenge in determining the criteria for separating the two varieties and, if parentage is unknown, for determining which variety is present. Additionally, as hobbyists are drawn to longer petals over shorter petals, breeders have responded by breeding var. roebelenii into the var. philippinense. Therefore, the average horizontal spread of var. philippinense has diminished over the last three decades. For example, philippinense ‘Penn Valley’ AM/AOS, awarded in 1980, had a horizontal spread of 6.3 inches (16 cm), and philippinense ‘Seven Samurai’ AM/ AOS, awarded in 2018, had a horizontal spread of 2.9 inches (7.4 cm).
Paphiopedilum philippinense has been in cultivation for many generations, and the cultivars used today for breeding are distant from wild-collected specimens. As a result, modern philippinense is much easier to bloom and grow. Using modern philippinense also makes its hybrids hardier. Paphiopedilum philippinense contributes flower count, narrow petals and a narrow dorsal sepal to its progeny. Breeders have tended to cross philippinense to other members of subgenus Paphiopedilum Section Coryopedilum (Cribb 1998), as they have similar characteristics in flower count, dorsal sepal width, and petal width. Paphiopedilum philippinense will always narrow the petals, which can be extremely detrimental to flower form when the other parent is a rounder Paphiopedilum such as a bulldog-type complex hybrid or members of the subgenus Parvisepalum (Cribb 1998). Paphiopedilum philippinense has a 2n chromosome count of 26. When breeding outside of its subgenus, the ploidy can be a mismatch and the fertility of the offspring goes down. In these cases, it is impossible to obtain a F2 generation. This can be seen in the lack of offspring from intersubgeneric crosses.
As stated previously, breeders such as Sam Tsui of Orchid Inn and Dave Sorokowsky of Paph Paradise are breeding philippinense var. philippinense and the other varieties of philippinense back to var. philippinense to create easierto-grow, faster-to-bloom, and more floriferous philippinense plants. They are also seeking to produce flowers with darker color, with longer petals or, to accommodate the increasing population of windowsill hobbyists, smaller plants. For example, Sam Tsui’s philippinense ‘Seven Samurai’, received an 83-point AM/AOS on its first blooming, with seven flowers on one inflorescence as well as an FCC of 91 for philippinense ‘Sam’s Choice’ as a first-bloom seedling.
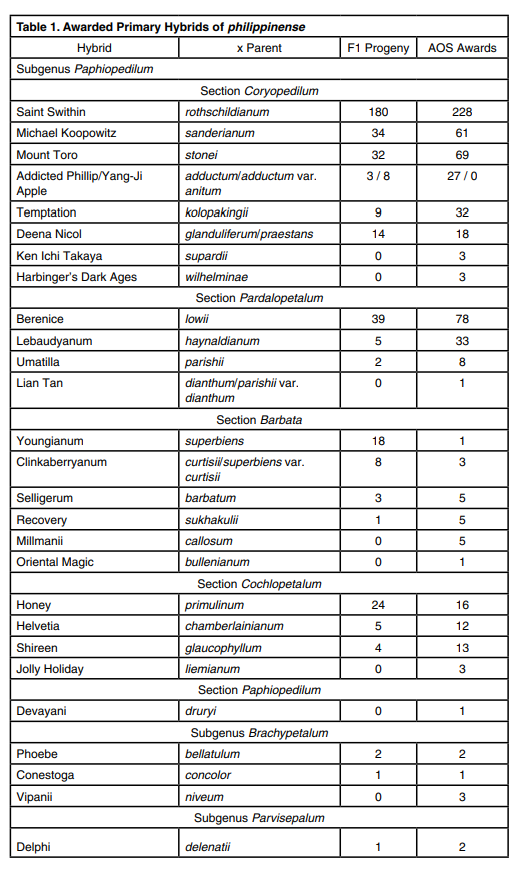
The most successful hybrids of philippinense are those made within its subgenus, especially within its section. This can be seen through the number of awarded plants and the number of second-generation or F2 progeny (Table 1). Though many primary hybrids have been made with philippinense, those made with paphiopedilums in subgenera Parvisepalum and Brachypetalum, based on a search of registered hybrids, have fewer F2 progeny than the primary hybrids made between philippinense and members of its own section. The four most popular and famous hybrids of philippinense are Paphiopedilum Saint Swithin (× rothschildianum), Paphiopedilum Berenice (× lowii), Paphiopedilum Michael Koopowitz (× sanderianum) and Paphiopedilum Mount Toro (× stonei). The parents of three out of four of these primary hybrids belong to subgenus Paphiopedilum Section Corypedilum, and one is from Section Pardopedilum (lowii), the section most closely related genetically through sequencing analysis (Chochai et al. 2012).
The success of Saint Swithin is unsurprising, given that Saint Swithin is the primary hybrid of philippinense and rothschildianum, registered in 1901. There are 228 AOS awards for Saint Swithin in OrchidPro. It is a registered parent to 180 second-generation philippinense progeny. In Saint Swithin, philippinense contributes its longer petals and its vigor, and rothschildianum, as the “King of Paphiopedilums,” contributes its large size. This results in a flower as large or larger than a rothschildianum with a broad dorsal sepal and long draping petals, which can go from seed to bloom far more quickly than the 10–15 years rothschildianum has been known to require. When using smaller philippinense, such as var. laevigatum or var. compactum, as a parent to reduce the size of the plant and increase the maturity rate, Dave Sorokowsky has been able to create a Saint Swithin that bloomed only three years out of flask.
The horizontal natural spread of Saint Swithin is also dependent upon the form of philippinense used as a parent. Use of var. roebelenii will result in a smaller horizontal natural spread than when var. philippinense is used. This can be seen when comparing Saint Swithin ‘Sunset Valley Orchids’ FCC/AOS with a natural horizontal spread of 7.5 inches (19 cm) to Saint Swithin ‘Krull’s Prince’ AM/AOS with a natural horizontal spread of 5.3 inches (13.5 cm). Often, use of var. roebelenii will also result in a greater vertical natural spread due to the the longer petals of the var. roebelenii parent. This can also be seen in the petal length and vertical natural spread of Saint Swithin ‘Krull’s Prince’ AM/AOS, 9 inches (22.8 cm) and 11.1 inches (28.2 cm), respectively, and Saint Swithin ‘Sunset Valley Orchids’ FCC/AOS has 7.6-inch (19.2-cm) petals and a vertical natural spread of only 9.6 inches (24.5 cm). Visually and statistically, it can be inferred that Saint Swithin ‘Krull’s Prince’ AM/AOS has philippinense var. roebelenii as a parent, because the petal angle on rothschildianum is constant.
Paphiopedilum Berenice (lowii × philippinense) is a perennial favorite. It was registered in 1891, making it older than Saint Swithin. There are 78 awards to Berenice listed in OrchidPro. Its pouch resembles lowii, which also contributes wider petals. Once again, petal length and dorsal sepal width comes from philippinense. Although the base of the dorsal sepal on philippinense is considered narrow, it does not reflex in the manner exhibited by the dorsal sepal of lowii. In this case, the flat dorsal of philippinense somewhat abates the dorsal reflexing contributed by lowii. Color is variable in Berenice and highly depends on whether var. philippinense (lighter), var. roebelenii (darker) or an albinistic form of philippinense was used. Paphiopedilum Berenice ‘Blumen Insel’, FCC/AOS scored 92 points, commended for its improved size, form and incredible color, was upgraded from an HCC/AOS of just 75 points back in 1995. With the new, darker philippinense cultivars now mature and reaching first bloom, it is possible that future Berenice will also exhibit deeper color. Crossing lowii f. aureum with an albinistic form of philippinense results in a pale, albalike Berenice with peach tones. In 2013, Berenice ‘Omaha’ was awarded an HCC/AOS of 77 points with a specific notation in the description that its parents were the aureum form of lowii and an alba form of philippinense. Using alba parents, as noted in the award description of Berenice ‘Ophelia’s Grace’ HCC/AOS, results in a pale-yellow flower.
[10] Paphiopedilum Saint Swithin ‘Sunset Valley Orchids’ FCC/AOS; exhibitor: Fred Clarke.

[11] Paphiopedilum Saint Swithin ‘Krull’s Prince’ AM/AOS; exhibitor: Krull-Smith.

Paphiopedilum Michael Koopowitz is a cross between philippinense and sanderianum. It was not registered until 1993, most likely because of the many decades that sanderianum was not in cultivation. Yet, it has 61 AOS awards and 47 awards given by other orchid or paphiopedilum societies, making it extremely successful despite its recent registration. Paphiopedilum sanderianum is known for its long, curling petals that can reach up to 39 inches (1 m) in length. As philippinense has shorter petals, Michael Koopowitz cannot reach the immense vertical natural spread of sanderianum, but it gains variety in the corkscrew spirals, both looser and tighter. Paphiopedilum philippinense also contributes its wider dorsal sepal, but this is ameliorated by the increase in dorsal sepal length, resulting in a similar cupping of Michael Koopowitz flower that is also seen in the flowers of sanderianum. Like many other strap-leaf paphiopedilums, sanderianum is also an extremely slow species to grow from flask. It can take upwards of eight years before its first blooming. By crossing with philippinense, vigor is imparted to Michael Koopowitz, reducing the long wait from flask to bloom.
Unlike lowii, no albinistic forms of sanderianum are currently known. Albinism is a recessive trait in philippinense and it is unlikely that use of an albino philippinense with a conventional form of sanderianum would produce an albinistic Michael Koopowitz. Instead, sibling crosses or selfings would be required of the F1 progeny, resulting in albinistic Michael Koopowitz in approximately 25% of the F2 offspring. Sam Tsui of Orchid Inn has been attempting to create an albino Michael Koopowitz by this method, and it is something to look for in the coming years.
[12] Paphiopedilum Berenice ‘Blumen Insel’ FCC/AOS; exhibitor: Dr. and Mrs. Henry Clay Frick.

[13] Paphiopedilum Berenice ‘Omaha’ HCC/ AOS; exhibitor: Joe and Karen Lankton.

[14] Paphiopedilum Berenice ‘Ophelia’s Grace’ HCC/AOS; exhibitor: Mary Ann Denver.

[15] Paphiopedilum Delphi ‘T.O.P. Pick’ HCC/AOS; exhibitor: Jack Schendowich.

Paphiopedilum Mount Toro (stonei × philippinense) rounds out the top-four primary hybrids. Paphiopedilum stonei tends to be even slower growing than sanderianum. Like rothschildianum, it is a reliable bloomer once it begins to bloom, but the time from flask to first bloom follows the trend of the other three strapleaf parents of the top-four philippinense primary hybrids. Once again, philippinense brings its vigor to Mount Toro by making it easier to grow and reducing the time from flask to first bloom, and stonei brings its prominent white dorsal. The first stonei f. alboflavum was awarded in 1999 by the Taiwan Paphiopedilum Society (as f. album). A cross with an albinisitic philippinense finally produced two awardable albino Mount Toro cultivars at the Taiwan International Orchid Show in 2016.
Other members of section Polyantha with albino forms are haynaldianum (f. album), glanduliferum, kolokapingii and dianthum (f. album). An albino primary hybrid has already been awarded for Paphiopedilum Lian Tan (dianthum × philippinense). For kolokapingii, an albino form of Paphiopedilum Temptation has been made, but not yet awarded, and for haynaldianum and glanduliferum, the albinistic forms of Paphiopedilum Lebaudyanum and Paphiopedilum Deena Nicol have yet to be seen.
Paphiopedilum philippinense has been crossed with species from every section of subgenus Paphiopedilum, but the majority of crosses from subgenera Paphiopedilum and Parvisepalum are not awarded and not have any registered progeny. The most successful hybrids are between philippinense and subsections Sigmatopetalum and Cochlopetalum and subgenus Brachypetalum. Parvisepalum, with its rounder flowers, often do not mix well aesthetically with the long, thin petals of philippinense. This can be seen in Delphi (delenatii × philippinense), which has only two low-point-score HCCs since its registration in 1983. In a similar manner, breeding philippinense with complex “bulldog” paphiopedilums has the same issue. When the goal is to breed toward large, round flowers, then philippinense, with its consistent contribution of thin petals, immediately forces the shape away from the ideal. The contribution of the multifloral nature of philippinense is often quickly lost in the F2 and F3 generations, along with its petal dimensions.
[16] Paphiopedilum Michael Koopowitz ‘Falcon’ FCC/AOS; exhibitor: ShinYi Orchids; photographer: Jea Shang Photography.

[17] Paphiopedilum Mount Toro ‘Snow Peak’ FCC/AOS; exhibitor: Jeff Morris.

[18] Paphiopedilum Mount Toro ‘Shin Yueh #8’; exhibitor: Shin Yueh Orchids.

[19] Paphiopedilum Lian Tan ‘Tadpole’ AM/AOS; exhibitor: George Titterton.

It is possible that the accounting of hybrids and awards for Paphiopedilum Youngianum, Paphiopedilum Clinkaberryanum, Paphiopedilum Umatilla and Paphiopedilum Lian Tan is incorrect, because of the reclassification of species. In the case of Youngianum and Clinkaberryanum, Paphiopedilum curtisii was considered a species in its own right, but in 1988 was recognized as a variety of Paphiopedilum superbiens. Prior to 2001, both curtisii and superbiens var. curtisii were recognized and awarded as independent species. After 2001, all curtisii have been awarded as superbiens var. curtisii. Because superbiens var. curtisii was preferred over superbiens var. superbiens for hybridization and the award to Youngianum was given in 1980, it is not entirely clear whether superbiens var. curtisii or var. superbiens was used to make the single-awarded Youngianum. A similar situation exists for Umatilla and Lian Tan, both hybrids of dianthum. Described by Tang and Wang in 1940 as a species in its own right, Karas and Saito reclassified it as a varietal form of parishii, Paphiopedilum parishii var. dianthum, in 1982. OrchidPro records awards for parishii var. dianthum starting in 1989 and dianthum starting in 1997. Until 2004, both synonyms are used concurrently in OrchidPro. The Australian Orchid Council recorded its awards for this species as dianthum, whereas the AOS was using parishii var. dianthum. Starting in 2006, the AOS also switched to dianthum, but because of this change in name, it is possible that some awarded Umatilla cultivars may have parishii var. dianthum as a parent. It might be possible to separate Umatilla parentage by floriferousness, because parishii should average 8–9 flowers per inflorescence, whereas dianthum averages 3–5 flowers, giving a geometric mean of 5–6 flowers. Paphiopedilum Umatilla ‘May’ HCC/AOS and ‘Great Hawaiian’ HCC/AOS were awarded in 1987 and 1997 with only two and four flowers, respectively, and these two cultivars look very similar to ‘Black Tie Weltz’ AM/AOS awarded in 1993, which had a higher flower count at six flowers and one bud on its inflorescence. Unfortunately, it is not possible to determine after the fact whether ‘May’ HCC/AOS and ‘Great Hawaiian’ HCC/ AOS should really be known as Lian Tan. Indeed, Lian Tan was not even registered until 2007 and may be a more successful hybrid than can be credited from its one recent award. It is possible, with the name changes, that many more Lian Tan will be awarded in the future.
[20] Paphiopedilum Youngianum ‘Bozo’ HCC/AOS; exhibitor: W.W. Wilson.

[21] Paphiopedilum Clinkaberryanum ‘Orchid Loft’ AM/AOS; exhibitor The Orchid Loft.

[22] Paphiopedilum Umatilla ‘May’ HCC/ AOS; exhibitor: W.W. Wilson.

[23] Paphiopedilum Umatilla ‘Great Hawaiian’ HCC/AOS; exhibitor: Great Hawaiian Orchids, Ltd.

[24] Paphiopedilum Umatilla ‘Black Tie Weltz’ AM/AOS; exhibitor: S.R. Weltz, Jr.

At least two primary hybrids, which have not reached the award numbers of the top four hybrids, should be considered up and coming: Temptation and Paphiopedilum Addicted Phillip. Paphiopedilum Temptation is (kolopakingii × philippinense). Paphiopedilum kolopakingii is very floriferous and can increase the flower count of its hybrids. This makes it a popular parent for breeders. The cultivar ‘Wacousta’ was awarded an FCC of 90 points in 2017 with six flowers on a single inflorescence. There have even been awarded plants with eight flowers on an inflorescence and reports of plants producing up to 15/inflorescence. However, it is a very large plant and its hybrids are not easily grown by hobbyists, because of the space necessary to keep them. Dave Sorokowsky has been crossing kolopakingii with the compact form of philippinense in attempts to have both floriferousness and a smaller plant with great vigor.
Paphiopedilum adductum var. anitum and Paphiopedilum anitum is another pair of names that spawn confusion. Paphiopedilum adductum was described by Asher in 1983. In 1998, a particularly dark form of adductum was described by Galamco as anitum and then reclassified in 2000 by Koopowitz as adductum var. anitum, where it is currently accepted. Paphiopedilum Addicted Phillip, registered in 1991, is (philippinense × adductum) and in this breeding line, the goal is to allow adductum to darken and intensify the color in the cross. With that stated goal, the question must be asked as to whether adductum var. anitum was the adductum parent or a more typically colored variant. The situation is further complicated by the fact that the Hybrid Register continued, until quite recently, to accept registrations using anitum as a parent, in effect creating darkly colored strains of prior registered grexes under new names.
[25] Paphiopedilum Addicted Phillip ‘Dark Passenger’ AM/AOS; exhibitor: Cheyenne Miller.

Paphiopedilum Yang-ji Apple is an example. It is commonly believed that all Addicted Phillip breeding and awarded plants were actually made with an anitum form of adductum, which is preferred by breeders for its darker color. However the cross of philippinense by adductum var. anitum is registered as Yang-Ji Apple. There are no AOS awards to Yang-Ji Apple compared to 27 awards for Addicted Phillip, but it is a popular parent for crosses made overseas and is a parent in eight registered grexes compared to the three plants naming Addicted Phillip as the parent. Comparison of the awarded Addicted Phillip ‘Dark Passenger’ AM/AOS to a Yang-Ji Apple owned by Tony Casler shows striking similarities. There are also many unregistered crosses with YangJi Apple, and some may begin showing up before judging teams as the imports become more popular.
Complex hybridization with strap-leaf paphiopedilums produces inconsistent results. Therefore, philippinense is often bred back to its own F1 progeny or crossed with other primary hybrids when complex hybridization is attempted. For example, it has been successfully crossed with Paphiopedilum Bel Royal, a primary hybrid between rothschildianum and kolopakingii, to create Paphiopedilum Crouching Tiger. This cross looks like a more floriferous version of Saint Swithin, and has already garnered 12 AOS awards, including an FCC, since its registration in 2008. Paphiopedilum Saint Swithin has also been crossed back to philippinense for a double dose of the species in Paphiopedilum Moustache. This hybrid has 32 AOS awards, including five FCCs, since its registration in 1982. Another successful hybrid, which mimics Saint Swithin in many aspects, is Paphiopedilum Kemp Tower (Prince Edward of York × philippinense); it looks like Saint Swithin but with longer petals. Paphiopedilum Kemp Tower has 15 AOS awards, including two FCCs. All three of these hybrids show that when rothschildianum combines philippinense, even with other paphiopedilums in the mix, the result is a differently colored or sized Saint Swithin. For reasons not yet well understood, the desirable traits of philippinense do not pass on to the majority of its progeny when it is more than one generation back, and often there are very few viable seedlings in the flask (Table 2).
Paphiopedilum philippinense has also been crossed with Maudiae-type paphiopedilums, most likely as an attempt to increase flower count in the latter. Maudiae-type paphiopedilums originate from hybrids of Paphiopedilum callosum and Paphiopedilum lawrenceanum, two species in subgenus Paph. Section Barbata. Maudiae-type paphiopedilums already have narrower petals and the further attenuation of the petals by philippinense will not be as detrimental to aesthetics. For the most part, the awarded hybrids have scored lower, and none have been awarded an FCC/ AOS. Both callosum and lawrenceanum typically produce a single flower, rarely two, per inflorescence. When crossed to philippinense making Paphiopedilum Millmanii and Paphiopedilum Charles Steimetz, respectively, the hybrids have 2–3 flowers/inflorescence. Neither of these hybrids have registered F1 progeny, nor does Paphiopedilum Weather Vane (Maudiae × philippinense), suggesting that an F2 generation is not viable. Therefore, it appears that the only way to have a Maudiae-type paphiopedilum with multiple flowers is to breed one with a multifloral as an F1.

Paphiopedilum Green Horizon was made by crossing the popular Paphiopedilum Makuli (Maudiae × sukhakulii) with philippinense. Paphiopedilum Makuli produces one flower/ inflorescence, occasionally two on a particularly well-grown plant. Awarded Green Horizon average 3.4 flowers/ inflorescence. The petals are not substantially longer and range from being the same width as the Makuli parent to at least twice as wide as the philippinense parent. The dorsal sepal also contracts in width, the influence of philippinense, but not in an unbalanced fashion, as the synsepal ends up more symmetrical. The awarded clones are all albinos, most likely made with an albinistic Makuli and one of the albino clones of philippinense. Use of a colored clone of Makuli results in a colored Green Horizon whose patterning is more similar to the Makuli parent than philippinense, suggesting that color in the cross comes from the Makuli side. All awarded Paphiopedilum Lime Twist and Paphiopedilum Somers Phil were made with albino Maudiae-type parents. Only Paphiopedilum Red Dragon involved a colored Maudiae-type parent. In all these crosses, flower count consistently runs 2–4 flowers/inflorescence, reinforcing the conclusion that philippinense only provides a boost in floriferousness, whereas other features, such as color and form, follow the Maudiae-type parent.
In summary, philippinense produces excellent primary hybrids when bred to other multifloral paphiopedilums. It is less successful as a parent outside Section Coryopedilum. There are many different forms of philippinense, providing a large genetic diversity, which allows for breeding in multiple directions at once. First and foremost, it contributes its vigor, creating easy-to-grow hybrids. Size reduction will make philippinense and its hybrids more accessible to hobbyists in colder climates or where space is a factor. Lovers of the long, curling elegance of the petals will enjoy the increase in floriferousness in the new cultivars. The horizontal natural spread of philippinense primary hybrids is greatly influenced by the choice of var. philippinense or var. roebelenii as the parent. Finally, breeders are choosing to breed for color in both directions: albino and saturated dark tones. With the shorter time required to go from flask to flowering of hybrids with vigorous philippinense as a parent, many of these hybrids will be appearing upon the scene in as little as five years and are something to anticipate.
[26] Paphiopedilum Crouching Tiger ‘Julio David’ FCC/AOS; exhibitor: Julio David Rios.

[27] Paphiopedilum Moustache ‘Marion Smith’ FCC/AOS; exhibitor: Krull-Smith.

[28] Paphiopedilum Kemp Tower ‘Joan’ FCC/AOS; exhibitor: Dr. and Mrs. Stephen Feairheller.

[29] Paphiopedilum Millmanii ‘Sentinel’s Ghost’ HCC/AOS; exhibitor: Mark A. Werther.

[30] Paphiopedilum Green Horizon ‘Joan’ HCC/AOS; exhibitor: Dr. and Mrs. Stephen Feairheller.

[31] Paphiopedilum Lime Twist ‘Judy’s Joy’ AM/AOS; exhibitor: Judy Cook.

[32] Paphiopedilum Red Dragon ‘Volcano’ HCC/AOS; exhibitor: Akatsuka Orchid Gardens.

REFERENCES
Chochai, A., I.J. Leitch, M.J. Ingrouille, and M.F. Fay. 2012. Molecular Phylogenetics of Paphiopedilum (Cypripedioideae; Orchidaceae) Based on Nuclear Ribosomal ITS and Plastid Sequences. Botanical Journal of the Linnean Society 170(2):176–196. doi: 10.1111/j.1095- 8339.2012.01293.x.
Cox, A.V., A.M. Pridgeon, V.A. Albert, and M.W. Chase. 1997. Phylogenetics of the Slipper Orchids (Cypripedioideae, Orchidaceae): Nuclear rDNA ITS Sequences. Plant Systematics and Evolution 208(3/4):197–223.
Cribb, P. 1987. The Genus Paphiopedilum. In: Christopher Grey-Wilson and Victoria Matthews, editors, A Kew Magazine Monograph. The Royal Botanic Gardens, Kew in association with Timber Press, Portland, Oregon. _. 1998. The Genus Paphiopedilum. In: Brian Mathew, editor, A Botanical Magazine Monograph: Natural History Publications (Borneo), 2nd ed. Royal Botanic Gardens, Kew
Gruss, O. 2016. “Paphiopedilum philippinense var. compactum f. aureoviride, Eine Neue Farbform. Die Orchidee 2(3):3–4.
Gruss, O., L. Roellke, and J. Roeth. 2008. Paphaphiopedilum philippinense var. compactum. Die Orchidee 59(3/4):316.
Hooker, J.D. 1865. Cypripedium laevigatum. Curtis’s Botanical Magazine 91, ser. 3, v.21:t5508
Karasawa, K., and K. Saito. 1982. A Revision of the Genus Paphiopedilum (Orchidaceae): Hiroshima Botanical Garden, Hiroshima, Japan.
Kent, A.H. 1889. Cypripedium philippinense. In J. Veitch & Sons A Manual of Orchidaceous Plants Cultivated Under Glass in Great Britain, PL 4:42.
Linden, J. 1887. Cypripedium cannartianum hort. Lindenia; Iconographie des Orchidees 3:Pl. CXLI, 93.
May 1885. Deux Nouveaux Selenipedium. Revue Horticole (Paris) 57:301–302
Pfitzer, E.H.H 1888. Paphiopedilum laevigatum. In H.G.A.Engler & K.A.E. Prantl, Die Natürlichen Pflanzenfamilien 2(6):84._. 1895. Paphiopedilum roebelenii. Botanische Jahrbücher fur Systematik, Pflanzengeschichte und Pflanzengeographie19:41.
Reichenbach, H.G. 1862. Trias Orchidacea Philippinensis. Bonplandia 22. _. 1883. Cypripedium Robbelenii. The Gardeners’ Chronicle: A Weekly Illustrated Journal of Horticulture and Allied Subjects n.s.:v.19.
Rolfe, R.A. 1912. Cordula philippinensis. Orchid Review 20:2.
Stein, B. 1892. Stein’s Orchideenbuch. Beschreibung, Abbildung und Kulturanweisung der empfehlenswertesten Arten. Mit 184 in den Text gedruckten Abbildungen. P. Parey, Berlin.
—Dr. Kay Perry started growing orchids during graduate school at Northwestern University. She received her PhD in biochemistry and biophysics in 2002 and performs basic biomedical research in collaboration with scientists from around the world investigating organisms such as the SARS-COV-2 coronavirus. In her free time, she applies the research techniques she learned in graduate school toward learning more about orchids. She is currently a student judge with the Chicago Judging Center (email spencer.aloysius@gmail.com).







